



1 引 言
早在1907年,Gates[4]就开始利用人工监测蜂群的温度变化用于指导蜜蜂养殖。在过去的一个世纪中,学者们通过多样的定量手段对蜂群的温度、湿度、重量、声音、蜜蜂的进出量、气体浓度以及振动等一系列特征进行了检测,并试图建立检测特征与蜂群活动之间联系,奠定了蜂群连续监测技术的理论基础[5]。在温度检测方面,相关研究表明,当蜜蜂的体温低于9~12℃时会发生昏厥,而适合蜜蜂幼虫生长的温度在35℃左右[6]。当环境温度过低时,蜂群会通过调节新陈代谢和收缩肌肉来产热;而当温度过高时,蜂群会通过扇风和增加蜂箱内的湿度来降温[7]。因此,蜂箱内的温度稳定性体现了蜂群对环境的适应能力和自身的健康状况[8]。同时,由于蜂群分蜂前的摇摆舞、集中决策等过程会造成蜂群的大量活动,导致蜂箱内的温度明显升高。因此温度的变化也可以作为分蜂等典型事件出现的依据[9,10]。在湿度检测方面,Doull[11]通过研究不同湿度下蜜蜂卵的孵化情况,发现最佳的孵化相对湿度为90%~95%,当相对湿度低于50%时,所有的蜜蜂卵都不能被孵化。然而,蜂箱内部湿度是多变的,它不仅受到外界湿度、蜂箱内的温度以及花蜜的含水量等因素的被动影响,同时也受到蜜蜂采水、喂养和身体隔离等活动的主动干预。因此,蜂箱内部的湿度变化较为复杂,而且不同的区域有较大的差异。Human等[12]采用多个传感器分别对周围环境,蜂箱育虫区和花蜜区的温湿度进行了监测,其结果证明幼虫区域具有最高的湿度和最好的稳定性,其相对湿度在40%左右。显然,他们监测结果明显的低于Doull等所提及的最佳范围,由此说明极端条件下湿度的控制是一个十分严峻的问题。在蜂群重量检测方面,Hambleton[13]采用每小时一次的人工称重方法,连续几天监测同一蜂箱的重量变化,发现蜂箱的重量变化具有很强的规律:夜间的蜂蜜浓缩和清晨的蜜蜂外出会造成蜂箱的质量减少,而上午到傍晚蜂箱的重量增加取决于蜜蜂的采收活动。因此蜂箱重量变化的幅度可作为蜂群日常活动的重要指标。进一步地,Meikle等[14]基于17个多月的连续重量监测结果,提议将蜂箱每七天的平均重量变化作为蜂群食物存量变化的依据,而每天的重量波动作为蜂群对食物消耗量的依据,因此通过这两个重量指标就能在不干扰蜜蜂的情况下揭示蜜蜂的活动和蜂群的发展。与此同时,由于蜂群发生分蜂和飞逃时,大量蜜蜂会携带蜂蜜离巢,也会造成重量瞬时的突变,因此一定幅度的重量突变可以作为分蜂和飞逃的判断依据。在声音检测方面,蜂群的声音是由众多蜜蜂个体活动所引起的空气振动的叠加。早在1957年,Frings和Little[15]就发现频率在300~1000 Hz并且具有足够强度的声音会使得蜂群中几乎全部蜜蜂的活动暂停。无论是意大利蜜蜂(Apis mellifera ligustica)、东方蜜蜂(Apis cerana)还是胡蜂(Vespa manderinia)都会产生特定频率的声音[16]。而在分蜂过程中,蜜蜂拍打翅膀的频率也存在着差异,获得分蜂指令的蜜蜂每隔0.5~3 s会扇动一次翅膀,其发出的声音频率在180~250 Hz之间,而当它们接触到其他蜜蜂时声音的频率会迅速提升至大约500 Hz[17]。因此,声音信号可以用来识别蜂群的活动,是一种具有应用潜力的检测特征。在进出量检测方面,由于蜂群中18日龄以上的蜜蜂负责外出觅食的工作,因此通过记录蜜蜂进出巢的次数就能够了解蜂群摄取食品的能力和种群的结构。为定量化地的分析蜂群的进出巢情况,一些人工[18,19]和自动[20-24]的计数方法被提出。Corbet等[19]发现蜂群存在一个明显的外出阈值温度,而且在一定温度范围内出巢的次数与温度成正比。同时,蜜蜂的进出数量也可以作为区分蜂群正常、中毒和分蜂等不同状态的依据[21]。蜂箱内的O2和CO2的浓度与蜜蜂的新陈代谢有关。Seeley[25]采用向蜂箱通入气体的方法,分别改变蜂箱内部的CO2、N2和O2的浓度,发现在巢门处扇风的蜜蜂数量随蜂箱内的CO2的浓度升高而正比增加。Southwick 和Moritz[26]观测到蜂箱内O2和CO2的浓度会像动物呼吸一样存在周期性的变化,其波动的幅值在0.6%左右,并且白天的波动频率为(2.9±0.84)次/min,是夜间的7倍。Nerum和Buelens[27]发现蜂群通过维持蜂箱内的低含氧量(<15%)来降低新陈代谢,从而度过食物匮乏的冬季。因此O2和CO2的浓度一定程度上可以作为判断蜂群健康状态的依据。蜜蜂扇风、摇摆舞等活动过程中都产生机械振动,而蜜蜂腿部也存在能够接收200~300 Hz振动的感受器。因此振动也是蜜蜂个体之间的一种重要的沟通方式,对信息的传播和行动的协调起到了关键性的作用[28]。特别是对于分蜂过程,Bencsik等[29]采用加速度计对蜂箱0~4000 Hz的振动信号进行了分析,发现500 和2000 Hz的幅值能有效的指示蜂群的分蜂行为。因此检测蜂群的振动信号有助于了解蜂群的内部活动[30,31]。
以上研究展示了蜂群活动与检测特征之间的密切联系。然而,蜜蜂是一种典型的社会性昆虫,它们的行为具有复杂多变的特点,通过单一的检测特征很难确定蜂群多样的行为,因此同时采用多种检测特征有助于提高蜂群活动的识别能力。Zacepins等[32]分析了温度、声音、出入和重量四种检测特征对蜂群的典型状态的识别能力。基于他们的分析,采用单群蜜蜂的数据可以对死亡、繁育、少子、预分蜂以及分蜂状态进行识别,而通过对比多群蜜蜂的数据可以进一步的识别蜂群的活跃程度、疾病、缺少食物以及异常进出。其中温度的识别能力最强,而其他的三种检测特征各有着不同的优点。由此也说明采用多种检测特征能够有效的提升识别的覆盖率和正确率。
随着电子信息技术的发展,信息化、智能化成为了农业发展的趋势[33,34],研究人员开展了系列基于连续监测的蜜蜂精确养殖[35-37]。尽管如此,国内外能够实现大规模、长时间实际监测的系统还十分缺乏,这是由于研制实用化的蜂群监测系统依然面临着巨大的挑战[32]。首先,蜂群是由成千上万只蜜蜂组成的复杂系统,采用单一传感器很容易发生蜂群状态识别错误,因此如何在系统中尽可能多地融合有用的传感器从而提高诊断的正确率是关键性问题。其次,监测系统能源是否自给决定了系统是否能够在野外长时间地工作。最后,因实际生产过程中需要频繁地搬运蜂箱,采用有线数据传输方式会带来极大不便,因此如何实现无线数据传输也是一个迫切需要解决的问题。
为定量化地监测蜂群行为和变化趋势,本研究在分析检测特征和蜂群活动之间关系的基础上,提出了一种基于太阳能供电的蜂群多特征无线监测系统,并将其应用于意大利蜜蜂蜂群实际监测中,实现了从2019年秋季到2020年春季为期235天的逐小时监测。最后基于监测数据分析了蜂群的活动。本研究的主要创新之处为:基于物联网技术设计了一种融合了多种传感器、太阳能电能自给和无线数据同步传输的蜂群监测系统,并实现了对意蜂蜂群多特征长时间的连续观测。
2 系统设计方案
2.1 特征适用程度分类
设计一套能被广泛使用的蜂群监测系统,除了考虑检测特征的多样性外,还需要考虑与蜂群的相关度、信息的丰富度、造价的经济性和实施的简易性等因素。因此,本研究依据文献和多年养蜂生产经验总结,评估了不同检测特征在蜂群监测系统中的适用程度,如表1 所示。通过不同特征之间的对比,将每种因素分为高、中、低三个量化等级。评估等级越高表明检测特征越适用。
表1 不同检测特征的适用程度Table 1 The adaptability of detection features |
| 检测特征 | 相关度 | 丰富度 | 经济性 | 简易性 |
|---|---|---|---|---|
| 重量 | 高 | 中 | 低 | 低 |
| 温度 | 高 | 中 | 高 | 高 |
| 湿度 | 低 | 低 | 高 | 高 |
| 气体浓度 | 低 | 中 | 低 | 中 |
| 振动 | 中 | 高 | 中 | 高 |
| 声音 | 高 | 高 | 高 | 高 |
| 进出量 | 高 | 高 | 低 | 低 |
这里相关度被作为最高的优先级进行考虑,重量、温度、声音和进出量四个检测特征与蜂群活动高度关联同时又具有不可替代性,因此被纳入监测系统的设计中。此外,因温湿度传感器为一体化设计,无需再为湿度检测的成本和技术实现付出更多的代价,尽管湿度的关联性和丰富度都为低,但也被采用作为检测特征。
2.2 监测系统设计
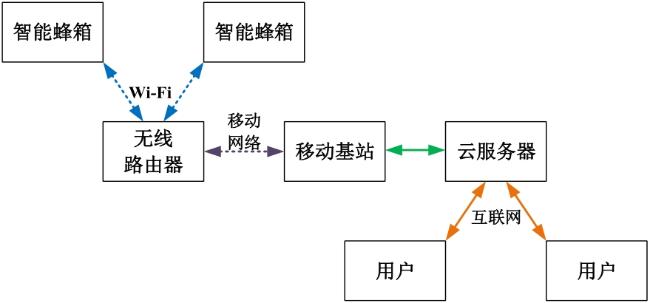
以往报道的蜂群监测系统通常以蜂场为单位进行内部数据存储与管理,这种方式存在标准不统一、数据无法共享、维护效率低等问题。随着物联网技术日趋成熟,通过标准的协议将监测对象的数据接入互联网,并集中存储于远程服务器中,已成为业内的共识。由于实际生产过程中,蜂箱会被放置在野外,同时根据季节变化可能会经常会被转运,因此利用无线网接入数据是最为理想的方式。手机移动网络是世界上覆盖最广泛的无线互联网接入方式,具有技术成熟、成本低廉的特点,可为蜂群监测系统提供数据接入方式。而由于每个蜂箱都需要传输各自的数据,如果各自单独接入手机网络,在功耗、成本和管理上都存在着劣势。因此本研究提出采用无线局域网(Wi-Fi)作为一个蜂场的内网进行数据通信,可以实现视距100 m的信号覆盖。如果以1 m的间隔部署蜂箱理论上可以实现上万箱的信号覆盖,能够满足实际部署的需求。基于以上考虑,研究构建了蜂群监测系统结构如图1 所示。每个智能蜂箱上布置一套用于采集蜂群信息的检测装置,并通过Wi-Fi上传至路由器,再经过移动网络上传至云服务器中,进行存储和分析。用户则可以通过访问云服务器了解蜂群的状态,从而进行蜂群管理。
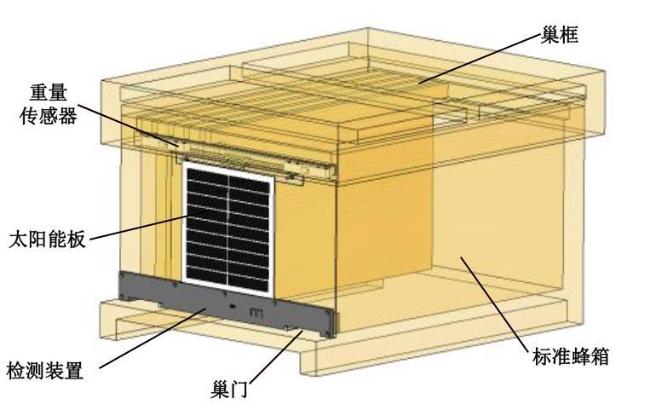
依据上述检测特征评估结果,本研究在蜂箱中安装了一套具有检测温湿度、重量、声音和蜜蜂进出量功能的检测装置,图2 是装有检测装置的智能蜂箱设计图。上述的五种传感器的安装位置为:温湿度传感器和声音传感器被集成在检测装置中,进出传感器安装在巢门处,称重传感器安装在蜂箱内部巢框的支撑面上。
2.3 检测装置技术方案
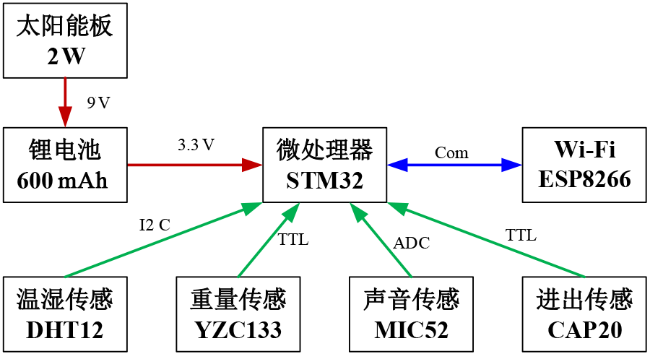
基于嵌入式方案能够有效地优化装置的功耗和布局,因此本研究采用嵌入式方案设计检测装置,图3 是技术方案原理图及参数选型。其中,太阳能电池板用于将太阳能转化为电能。锂电池作为电能的存储装置,当太阳能充足时将存储多余的电能,反之释放存储的电能以支持装置运行。Wi-Fi模块用于装置与服务器之间的数据通信,由于是成熟的商用模块,并基于TCP/IP协议具有通讯校验和重发机制,因此能够保证数据传输的可靠性。五种传感器分别用于获取各种检测特征数据。微处理器用于控制这些组件的协同工作。选用基于ARM内核的32位嵌入式处理器STM32作为装置的微处理器。选用基于AT指令集的ESP8266作为Wi-Fi通讯模块。此外,在本设计中,综合考虑量程和精度等因素,温湿度传感器采用基于I2C数字接口的DHT12模块,重量传感器采用成熟的电桥式称重单元YZC133,声音传感器采用MIC52由STM32的片上AD通道进行模数转化,进出巢量采用CAP20感应式模块进行检测以TTL电平方式与STM32进行连接。
为实现能源自给,需要尽可能地降低系统功耗,除了采用低功耗的元部件外,检测装置需要采用间歇工作模式来进一步降低整体功耗。同时,还需要保证太阳能的年度获取总量大于装置的年度消耗总量,以及具备足够大的锂电池能够支撑装置度过阳光匮乏季节。目前所设计的装置能够以每小时一次的频率将蜂群的信息传输至云服务器,并且经测试,能在没有任何阳光的情况下稳定运行两周以上。智能蜂箱的有关参数如表2 所示。
表2 智能蜂箱的参数Table 2 The parameters of smart hive |
| 名称 | 参数 |
|---|---|
| 箱体尺寸 | 内部37 cm×47 cm×26 cm |
| 巢口尺寸 | 60 mm×7 mm,2个 |
| 供电方式 | 锂电池+太阳供电 |
| 通讯方式 | Wi-Fi |
| 温度检测 | (-20~60℃)±0.5℃ |
| 湿度检测 | (20%~95%)±5% |
| 称重检测 | (0~20 kg)±2% |
| 进出检测 | 0~10,000只/h |
| 声音检测 | >-40 dB |
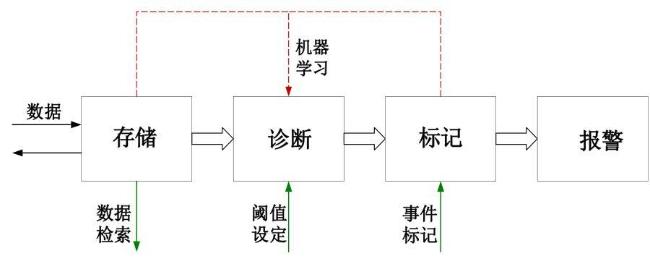
2.4 云服务器设计
3 系统监测试验和结果分析
3.1 试验方案
为了验证监测系统的性能,将研制好的智能蜂箱部署于山东省泰安市山东农业大学南校区,用于对若干群意蜂的活动进行监测。部署方式为传统的并排式放置,间距大约为10 cm,如图5 所示。

监测时间从2018年8月17日持续至2020年4月16日,监测蜂群编号为8321。
3.2 监测结果
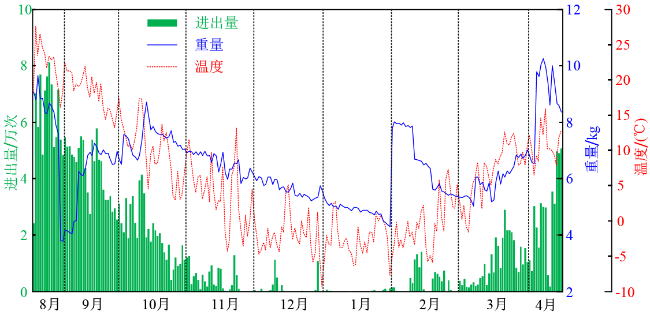
为进一步分析蜂群的日常活动,在蜂群作业比较有代表性的三个时期(采蜜、越冬、冬繁),各随机选取一天,分别是2019年8月25日、2020年1月15日和2020年4月16日,作为这三个时期的代表日期,如图7 ~9 所示。其中绿色柱状图为蜜蜂每小时通过巢门的累计次数,蓝色实线为每时刻蜂群的总重量,红色虚线表示每时刻蜂箱内的温度,紫色点划线表示每时刻蜂箱内的湿度。
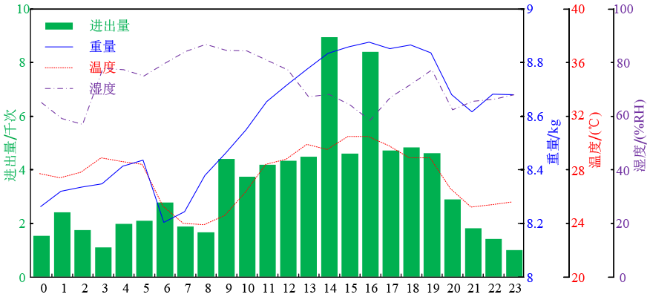
2019年8月25日为秋衰期中进出巢数量最大的一天,全天累计进出巢的次数达8万次。在这一天中,蜂箱内的温度变化在24℃~32℃,最低温度出现在上午的8~9点之间,最高温度出现在15~16点之间;蜂箱内的湿度变化范围为60%~80%,湿度的变化与温度呈现一定的负相关特性;由于蜂箱内的温度较高,蜂群夜间也存在进出巢的行为,全天进出巢主要集中在9~19点之间,特别是14点和16点存在两个高峰,这一现象可能与新蜂练飞相关;蜂群的总重量在这一天中增加了大约400 g,质量增加过程集中在6~16点之间,由此说明早上,特别是清晨对于蜂群采收蜂蜜至关重要,因此应该尽量避免在这一时间段人为的干扰蜜蜂的活动。
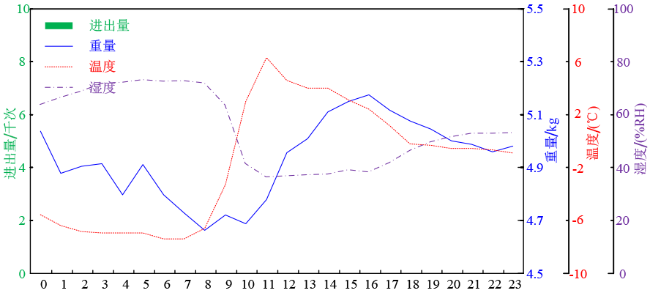
2020年1月15日是越冬期中的一天。期间,蜂群完全没有出巢行为。其蜂箱内的温度很低,早上6~7点甚至到达了-6℃,最高温度出现在11点为6℃,这一温度升高很有可能是由于阳光直射蜂箱引起的,由此也说明越冬期间阳光直射蜂箱有助于蜂群增强蜂群的越冬能力。这一天中,蜂群的总重量也存在着一定的波动,这一现象有可能与称重传感器的温度特性有关,也有可能与蜂群在蜂群上的活动有关。尽管如此,扣除总重量的波动仍然能够发现其总体呈下降的趋势。
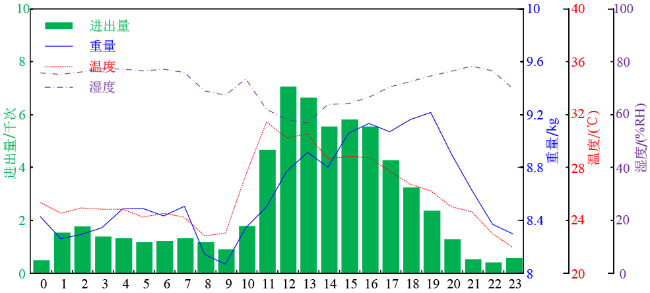
2020年4月16日是春繁期中进出巢次数最多的一天,累计达5万次。其监测结果与2019年8月25日相似,主要区别在于日间采收活动直到上午9点才开始而且时长较短。由于繁育期饲料消耗较大,尽管日间能够采收到花蜜和花粉,但是蜂群的总重量并没有增加,这也导致了春繁期蜂群的总重量波动较大。
进一步分析这三天蜂群的声音频谱,如图10 ~12 所示。
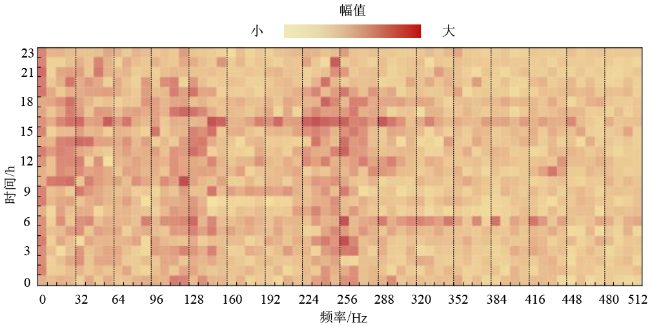
图10 蜂群2019年8月25日的声音频谱图Fig. 10 The acoustic spectrogram of bee colony on August 25th, 2019 |
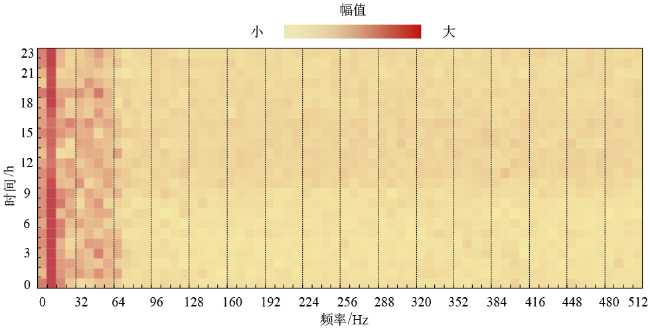
图11 蜂群2020年1月15日的声音频谱图Fig. 11 The acoustic spectrogram of bee colony on January 15th, 2020 |
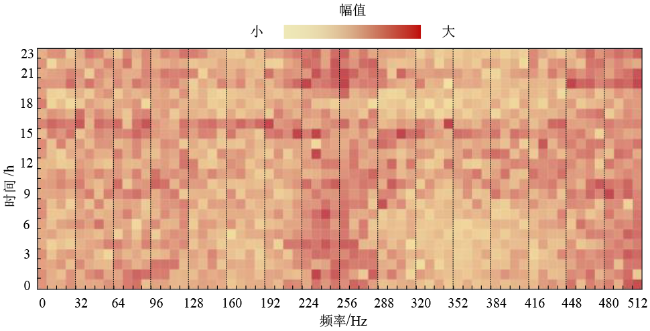
横坐标表示声音的频率,分辨率为8 Hz;纵坐标表示时间,单位为h;图像的颜色表示该频率声音的幅值大小。对比这三天的结果,可以发现春繁期蜂箱内的声音最大,秋衰期其次,越冬期最小。春繁期和秋衰期的频谱图具有一定的相似性:声音中均包含128 Hz和256 Hz附近的频率成分,并且全天持续存在。相关研究表明这两个频段的声音与蜜蜂的扇风活动有关。越冬时的频谱图与其他两个时期差异较大:几乎不存在大于64 Hz的频率成分,而小于64 Hz的频率信号持续保持相等的强度,由此推断小于64 Hz的声音频率信号与蜂群越冬过程中的产热机理有关,值得后续再开展深入研究。
4 结论与展望
本研究在分析蜂群行为与检测特征的基础上,设计了一种蜂群监测系统,它能够检测蜂箱内部的温度、湿度、蜂群重量、声音和蜜蜂的进出量七个特征,并通过手机移动网络和Wi-Fi将这些数据上传到远程云服务器中。相对于其他监测系统,该系统采用多传感器融合设计,能够实现电能自给和无线数据同步传输,方便系统部署和实现蜂群长期监测。
同时,基于监测系统,本研究开展了针对意蜂从秋季到春季的连续监测试验并分析了监测数据。数据反映了蜂群在秋衰期、越冬期和春繁期蜂箱内部温度、湿度、蜂群重量、声音和进出量的逐小时的细致变化。结果表明,该系统能够在不干扰蜂群活动的前提下揭示蜂群的日常活动和群势变化。其中,温湿度数据可以反映蜂箱内部的微环境状态,能够用于评估蜂群生存环境;蜂群的总重量变化和蜜蜂进出巢的次数可以反映蜂群的日常活动,能够用于评估蜂群的发展趋势;蜂群的声音反映了蜜蜂之间的协同活动,是一个值得进一步研究的特征,它将有助于增进对蜂群微观行为的了解。基于这种蜂群监测系统的优异性能,本系统可用来研究蜂群的行为生物学、探索崩溃式的蜂群消失成因以及促进发展精确化的蜜蜂养殖业。
后续将研究蜂群行为特征的建模,进而结合监测数据对蜂群的状态进行识别。






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}