



1 引 言
作物模型可分为描述性模型(Descriptive Model)和解释性模型(Explanatory Model)。前者又称为统计模型、回归模型、经验模型或黑箱模型,直接建立输入与输出之间的关系,不关心内在形成机理。后者又称为基于过程的模型(Process-Based Model),是指能定量地和动态地描述作物生长、发育和产量形成过程及其对环境反应的计算机模拟程序[6],通常是在群体水平对作物生长的描述。常见的基于过程的作物模型,如CERES[7]、TomSim[8]、STICS(Simulateur mulTIdisciplinaire pour les Cultures Standard)[9]和APSIM(Agricultural Production Systems sIMulator)[10]等,模拟作物的光合作用过程、同化物的分配过程等,主要通过叶面积指数(Leaf Area Index,LAI)来预测每平方米的生物量产量,同时考虑辐射、温度和基本投入(灌溉和化肥等)的影响。这类模型大多将不同类型的器官作为一个整体考虑,关注作物的总果实重量、叶重等,难以描述作物个体内结构的变化及其对产量的影响。以作物模型为核心的农业技术转移支持决策系统(Decision Support System for Agrotechnology Transfer,DSSAT)以天、日为步长模拟作物生长,按照作物生育期详细描述作物生长发育过程,包括发芽到开花、叶片出现、开花时期、籽粒灌浆、生理成熟和收获等,并且可响应许多因素,包括作物遗传特性、管理措施、环境、氮素和水分的胁迫、病虫害等,主要用于农业试验分析、农业产量预报、农业生产风险评估、气候对农业的影响评价等[2]。
由于作物结构的重要性,衍生了个体尺度的植物功能结构模型(Functional-Structural Plant Model,FSPM)来模拟作物形态结构、生物量的产生和分配以及两者的内在联系。FSPM是在单个器官到植物个体层次对植物这一复杂系统的建模与仿真[11],在过去的二十多年间得到了迅速的发展,成为植物建模的研究热点之一。
GreenLab模型是作物功能结构模型之一,考虑同类器官顺序产生的特点,并采用公共池和源库关系的概念,既在叶元尺度上(器官)对植物的生长进行建模,同时又保持了与基于过程的作物模型(群体)的兼容性。GreenLab模型基于植物自动机(Automaton)[12],通过递归算法计算每个时间步长产生的器官数量,再根据相对库强(Relative Sink Strength)和器官数量,将从公共池中获取的生物量分配到不同类别的器官中。该过程通过数学公式来描述,不需要逐个器官地来模拟生物量的分配,因此计算速度快,所需时间少,这是GreenLab模型的优势之一。此外,GreenLab模型的特色在于通过实测植物各个器官的重量等数据可反求影响生物量产生和分配的模型源库参数,已针对玉米[13]、小麦[14]、油菜[15]、黄瓜[16]、番茄[17]等十几种作物进行模型应用研究[18]。但是,GreenLab模型将环境因素的影响简化为一个环境因子E来描述,不能很好地模拟气候、土壤以及管理措施等对作物产量的作用。
近年来,随着作物表型信息获取设备的迅速发展,已实现采集作物株高、叶面积、籽粒数,以及生理和光合等各种表型信息。因此,如何对获取的大量表型数据进行分析是当前正待解决的问题[19]。不同作物模型的融合,有助于更深入地理解作物行为,更大程度发挥模型作用。本研究采用DSSAT系统与GreenLab模型进行群体尺度和个体尺度模型的融合研究,探索以不同空间尺度和方法所构建的模型进行交互的可能性,使得既可以模拟作物遗传特性、管理措施、环境等的影响,同时也可以模拟植物个体内各器官生长和发育的动态过程,根据对植物的更深层次的了解来详细描述产量的组成,并对作物进行三维展示,与表型信息做对比。通过两类模型的融合,为植物育种、表型鉴定和作物系统优化提供参考。
2 材料与方法
本研究总体思路如图2 所示。模型融合的思想考虑到发展模型的人员、模型实现方法的不同,且自成体系,选择以统一的输出结果进行连接,而不是尝试在模型构成模块进行融合。首先,利用DSSAT系统输出模拟的每日玉米的叶重、茎重和玉米棒重等;再基于这些数据,结合GreenLab模型已有的玉米的结构信息与库强变异参数等结果,计算模型的源库参数,输出不同叶位、不同生长阶段的各器官的详细数据,以及玉米植株的三维结构图。
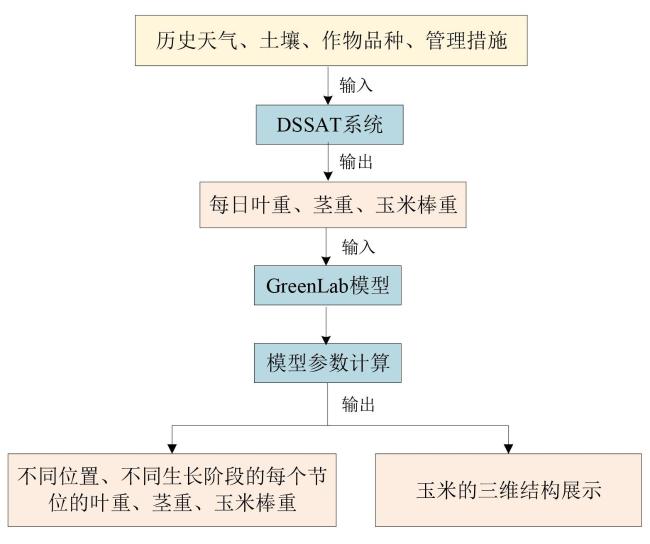
2.1 数据选取与处理
2.1.1 试验数据选取
试验数据包括两部分。(1)Soler等[20]2002年在巴西皮拉西卡巴(西经22.7°,南纬47.4°,海拔580 mm)进行的四个玉米品种(AG9010、DKB333B、DASCO32和EXCELER)两个处理,即灌溉(Irrigated,I)与雨养(Rained,R)的田间试验测量数据,包括土壤、气候、管理措施等,以及七次定期(3月12日、3月21日、3月27日、4月7日、4月18日、4月28日和5月14日)测量的玉米LAI和地上部总重量,共包含8个对比实验。其中,AG9010为短季品种(从种植到吐丝期为904生长度日(Growing Degree-Day,GDD),基温为8 °C),DASCO32与EXCELER为两个短季品种(995 GDD),DKB333B为常规品种(1037 GDD)[21,22]。所有试验为4个重复的随机区组设计。每个地块长20 m,行距0.8 m,共四行,种植密度为5 株/m2;(2)基于(1)中的数据,利用DSSAT系统模拟4个玉米品种的每日数据,包括每日叶、节、玉米棒和地上部总重等。本研究选取了叶片数分别为7、10、15、20、23、27、30、33、36、39和40共11个生长阶段的DSSAT系统模拟数据用于GreenLab模型的模拟计算。
基于(2)中的DSSAT系统模拟的每日数据拟合GreenLab模型的参数,模拟LAI、玉米叶、节和玉米棒的重量,以及地上部总重量,并比较四个玉米品种和两个处理的DSSAT系统和GreenLab模型模拟的数据与实验测量数据之间的差异性,分析模型结合的可行性。
2.1.2 数据处理
DSSAT模型以日为单位进行模拟,而GreenLab模型以叶元为单位进行模拟,每长出一片叶子即被认为是一个生长周期(Growth Cycle,GC),即出叶间隔(phyllochron),进行模拟。因此,需要将日转换为生长周期。通过将日历时间(天)转换为积温(温度总和)来调节植物的发育,从而可以定义周期的概念。
在GreenLab模型中,一个生长周期中一个分生组织产生一个叶元,叶子接收与该周期中累积的光辐射成比例的能量。根据DSSAT中的每日数据与积温,计算出GreenLab模型中对应的生长周期。根据种植密度计算出每株植物各类器官的重量。
为同步植物功能结构模型中的生长和发育,GreenLab中的生长模型采用与发育模型相同的时间尺度,即生长周期,从而简化了模型的计算。在每个周期中,分生组织产生新的叶元(发育);同时,叶片进行光合作用产生生物量并根据各自的库强分配到植株的各个器官中(生长)。因此,基于同一时间尺度模拟植物的发育和生长。这种方法使得当植物的顶端发育停止(雄穗出现)时,仍然可以确定其时间尺度。生长周期数与积温之间的线性关系是在植物发育阶段建立的,然后外推到植物生长阶段。
此外,DSSAT模型中的重量单位为g/m2,需要根据种植密度计算出每株植物的各类器官的重量,本研究主要考虑叶、节和果的重量。
2.2 模型参数设置
基于过程的作物模型原理是在日历(天)的每个时间步长计算单位面积土地截获的光所产生的生物量。植物吸收的光辐射量可以根据Beer-Lambert定律计算,公式为:
其中,k为消光系数;I 为冠层深度为L时的辐射水平,W/m2;I 0分别为冠层最上方的辐射水平,W/m2;k值一般在0.5至0.8的范围内[1]。光吸收量随叶片面积的增加而增加,但叶片遮挡会减少光的截获。
生物量可通过公式(2) 计算。
其中,Q(t) 为在时间t产生的生物量,g;RUE表示光能利用效率(Radiation Use Efficiency);LAI(t)是指在时间t测量的群体叶面积指数。通过对时间进行求和,可以通过累加计算总生物量[4]。
GreenLab模型是基于生物量公共池[23]以及源库关系假设的模型,并使用了作物生长年龄和生理年龄的概念。作物生长年龄指植物所经历的生长周期数,所有器官都有生长年龄。作物生理年龄用来区分植物中不同类型的枝,主茎生理年龄为1,分枝生理年龄为2。
GreenLab模型中的时间步长转化为生长周期表示。计算生物量生产的公式为:
其中,E为潜在光合生产能力,即将环境因素的影响简化为一个参数来表示;t为植物的生长年龄,即植株的生长周期,GC;φ为生理年龄,从1到mxφ,玉米为单茎植物;i,j为不同器官的生长年龄,GC;t a为叶子的功能周期,可根据经验或观测值设定;S p为植株投影面积,m2,取值为种植密度的倒数[13];ε为叶片厚度,一般为常数,可通过测量的叶片重量除以叶面积计算,g/cm2;r为光利用系数;k为Beer-Lambert定律的阻力系数,一般为0.7[13]; 为周期i时的叶子数,用于计算植株总的叶片面积; 为功能叶片的库强;D(j)为周期j的植物总需求; 为上一周期产生的植物总生物量,g;Q(0)为种子生物量,g。
为了计算植物的叶面积,需要将功能性叶子的重量相加,再除以它们的厚度ε(假定为常数)。可以通过公式(4) 计算功能叶面积Sf(t)。
其中, t为植物生长年龄,GC;i为叶片生长年龄,GC; 为植物生长年龄为t,叶片生长年龄为i时叶片的数量,个; 为植物生长年龄为t,叶片生长年龄为i时叶片的生物量,g。在生长年龄为i的一组叶片中,记录了叶片的数量 及其各自的生物量 。根据植物的年龄t是否超过叶片t a的寿命,需要划分为两种情况来计算面积。实际上,只要没有叶子死亡,t < t a的情况就相当于植物生长的开始。在另一种情况t ≥ t a下,只有生长年龄小于t - t a + 1的叶子仍然起作用,这部分通过公式(4) 中的第二个和的索引计算。
由于玉米生长后期叶和节的生物量会转移到果实中,因此,还需要考虑物质再分配过程(Remobilization),即成熟的器官(叶和节)重新分配部分生物量给果,计算公式如下。
其中,t为植物的生长年龄,GC;q maxo为该器官的最大重量, g;k mo为该器官转移的生物量的比例,%,可通过测量数据进行估计;k ko为生物量转移速度,g/GC,由模型反求得到;o表示器官类型,本研究中为叶b和节i。
2.2.1 模型生长参数设置
根据种植密度d 为5 株/m2设定S p的初始值为 1/d ≈ 2000,实际根据拟合结果进行调整。根据DSSAT中的光能利用效率(RUE:4.2 g/MJ) 以及GreenLab模型中的经验值设定与DSSAT中相对应的r值为50;根据DSSAT模拟的数据,设定叶、节和果的扩展周期分别为15、25和20,功能周期都为25,以及用于计算库强变异函数的参数[13];设定叶片的相对库强为1。
2.2.2 发育参数设置
对于玉米,每个叶元包含一个叶子、一个节或果。根据DSSAT中的数据,最终的叶子数为19~21个;根据DSSAT模型中的数据,四个玉米品种都为一个果的类型,位于主茎的第16或17个节上。
2.3 模型参数拟合
根据公式(3) ,将其中的r值固定,拟合环境参数E和库强参数P o(o为叶子b、节i和果f),S p的值根据模拟结果进行适当调整。根据经验,环境的变化对GreenLab模型中的库强参数影响不大[13]。因此,尝试采用一个环境参数的平均值E,而不是读取环境序列进行模拟。此外,还需要拟合物质再分配(Remobilization)的两个参数k m和k k。其中,转移比例k mb和k mi根据实验数据估算得到,均为0.8;转移速度参数k kb和k ki由模型拟合得到。
3 试验结果与分析
3.1 试验结果
3.1.1 株高和单位面积最终产量
四个玉米品种的两个处理收获时的株高和单位面积最终产量数据如表1 所示。灌溉处理的株高和单位面积最终产量均高于雨养处理的植株。其中,品种EXCELER两个处理的产量均最大,品种DASCO32次之,品种AG9010最小。结果表明,灌溉可以显著提高四个玉米品种的产量,且EXCELER是最优品种。
表1 四个玉米品种两个处理的株高和单位面积最终产量Table 1 Plant heights and final yield of the two treatments for the four maize cultivars |
| 品种 | 株高/cm | 产量/(g·m-2) | ||
|---|---|---|---|---|
| 灌溉 | 雨养 | 灌溉 | 雨养 | |
| AG9010 | 200 | 175 | 498.6 | 404.4 |
| DASCO32 | 217 | 206 | 504.7 | 410.9 |
| EXCELER | 230 | 227 | 530.6 | 485.9 |
| DKB333B | 213 | 202 | 513.9 | 382.2 |
3.1.2 DSSAT模拟的各类器官重量
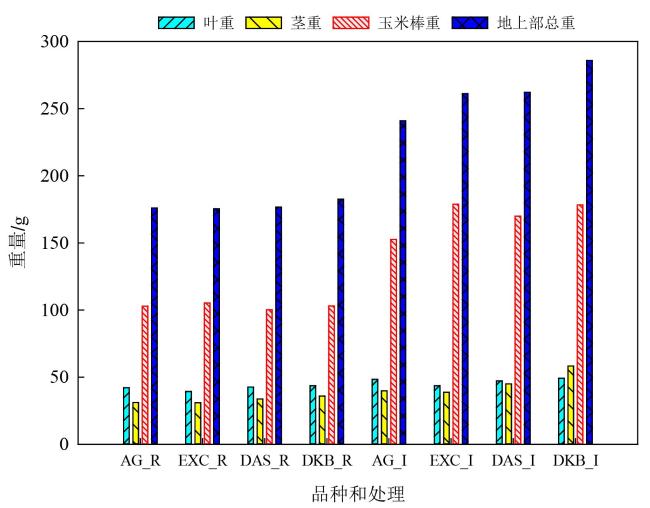
四个玉米品种的两个处理收获时的叶子、节、果各类器官重量如图3 所示。灌溉处理的各类器官重量均高于不灌溉处理。但DSSAT模拟的DKB333B的产量最大,与试验测量结果偏差较大。DASCO32和EXCELER的产量相近,AG9010的产量最小,与表1 试验结果一致。
3.2 模型模拟结果
3.2.1 两个模型时间尺度的转换
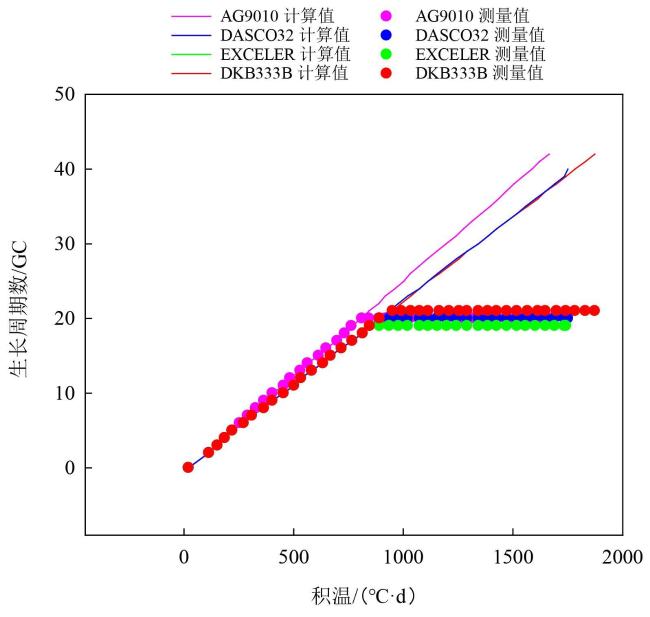
四个玉米品种灌溉和雨养处理的叶子数和理论生长周期数随积温的变化相同(图4 )。因此,只给出了灌溉处理的四个玉米品种的生长周期随积温的变化。不同品种间的叶子数无显著差异(19~21),且三个短季品种AG9010、DAS CO32与EXCELER所需要的生长周期数无差异,仅常规品种DKB 333B的生长周期数略长,但差异不显著(F = 0.097,P > 0.5,df = 3)。
根据温度和叶片数量可以计算出叶间隔。对于玉米,积温与所发育的叶元数量之间呈线性关系。因此,可以计算出以积温表示的生长周期,以及各生长周期对应的叶元数量。
当叶片数量不再增加后,玉米到达成熟期,该阶段虽然不再产生新的叶元,但生长仍在继续。可以根据从发芽到植物顶端抽穗阶段的积温和叶片数量之间的二元线性关系(斜率)来确定理论的生长周期数。即将时间离散化,根据发育阶段确定的线性关系来计算生长阶段的理论生长周期数(图4 )。
3.2.2 模型源库参数
对于每个实验,通过将参数S p、r,以及库强变异参数设置为固定值,可以拟合环境参数E、库强参数(P i和 P f)以及物质再分配参数k kb和k ki。
拟合的参数值如表2 所示,可以看出四个玉米品种灌溉处理的E、P i、P f和S p的值均大于不灌溉处理。其中,EXCELER两个处理的果实库强(P f)最大,DASCO32和DKB333B次之,AG9010最小。该结果与试验结果一致(见表1 ),四个玉米品种两个处理的模拟值与测量值的相关系数均为0.99,表明模型的模拟精度较高。
表2 四个玉米品种两个处理的拟合参数值Table 2 Parameter values of the two treatments for the four maize cultivars |
| 品种 | 处理 | E | P i | P f | k kb | k ki/(g·GC-1) | S p/m2 |
|---|---|---|---|---|---|---|---|
| AG9010 | 灌溉 | 0.17 | 0.96 | 243.3 | 0.014 | 0.030 | 3000 |
| 雨养 | 0.15 | 0.76 | 71.9 | 0.016 | 0.035 | 2500 | |
| DASCO32 | 灌溉 | 0.23 | 1.26 | 692.6 | 0.014 | 0.030 | 2200 |
| 雨养 | 0.17 | 0.89 | 112.9 | 0.013 | 0.033 | 2150 | |
| EXCELER | 灌溉 | 0.21 | 1.29 | 865.8 | 0.015 | 0.035 | 2500 |
| 雨养 | 0.17 | 1.03 | 143.2 | 0.015 | 0.050 | 2200 | |
| DKB333B | 灌溉 | 0.21 | 1.22 | 577.6 | 0.015 | 0.030 | 2100 |
| 雨养 | 0.18 | 0.94 | 114.9 | 0.016 | 0.032 | 2000 |
3.3 两个模型模拟结果与测量结果比较
由于田间实验只测量了LAI和地上部总重量,因此,将两个模型的LAI和地上部总重量的模拟结果与实验数据进行对比,以验证模型的准确性。
3.3.1 LAI的比较
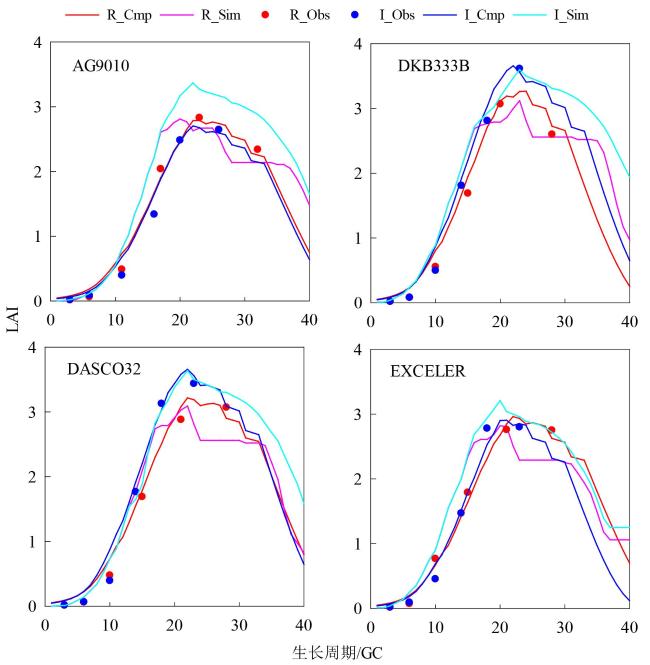
3.3.2 地上部总重量的比较
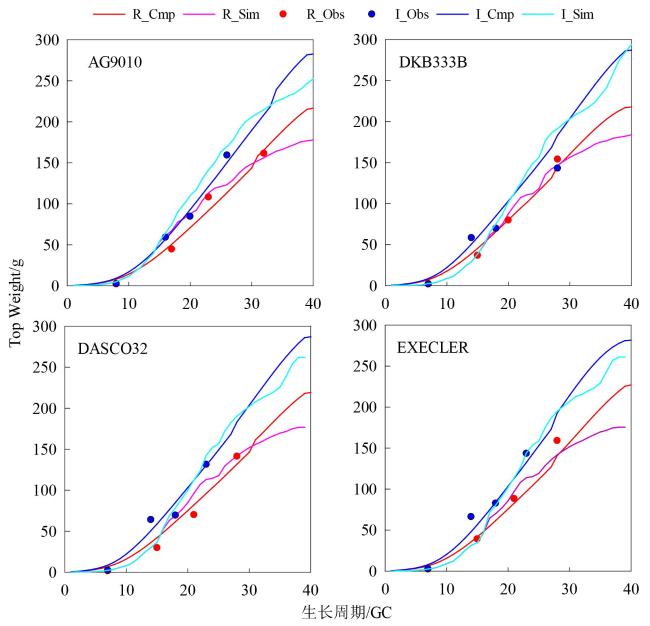
3.4 GreenLab与DSSAT模拟结果比较
3.4.1 不同生长阶段各类器官重量模拟
GreenLab模拟的四个玉米品种两个处理的不同生长阶段各类器官重量结果均非常好,所有模拟的R 2均为0.99,如图7 所示。结果表明GreenLab模型可以很好地计算这些值,计算结果与DSSAT的模拟结果一致。

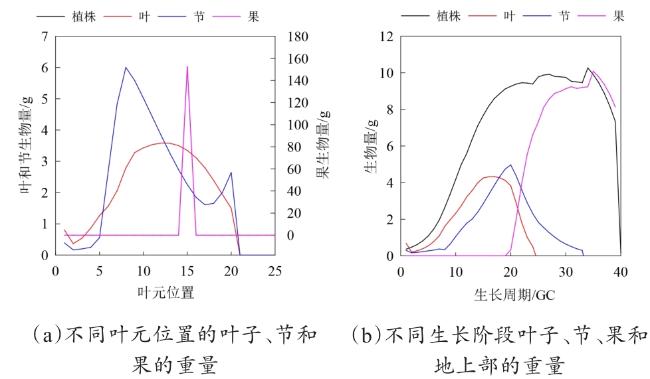
3.4.2 各类器官重量随位置和生长阶段的变化模拟
GreenLab模型是基于器官尺度的模型,可以模拟各类器官重量随位置(图8(a) )和生长阶段(图8(b) )的变化情况。叶子和节的重量随叶位和生长阶段均呈先增加后减小的趋势。
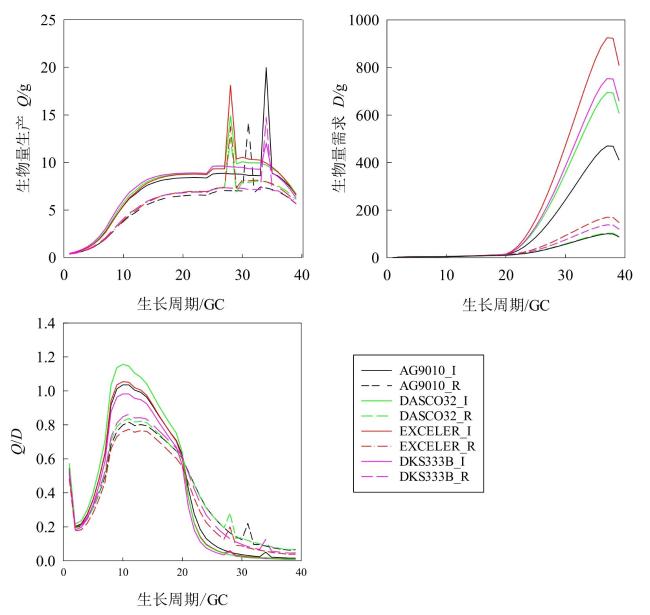
3.4.3 生物量生产和需求的变化模拟
GreenLab模型可以模拟植物生长过程中的生物量生产Q和需求D的变化情况。四个玉米品种灌溉处理的Q和D值均大于雨养处理,而发育开始时灌溉处理的Q/D比均大于雨养处理,但叶片停止发育后,灌溉处理的Q/D比均小于雨养处理(图9 )。
3.4.4 玉米植株的三维结构模拟结果
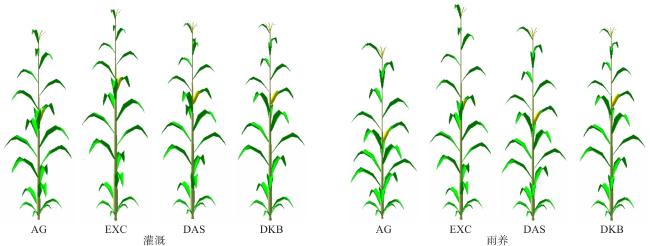
4 讨 论
4.1 不同尺度模型连接的可行性
本研究基于DSSAT的模拟数据,利用GreenLab模型可以很好地模拟出LAI和地上部总重量,与试验结果一致。此外,GreenLab模型计算的不同阶段各类器官(叶片、节、果)的总重量也与DSSAT的模拟数据一致。由此可见,DSSAT与GreenLab之间具有良好的兼容性。初步探讨了连接传统的作物模型和功能结构模型的方式[26]。尽管不一定改进基于过程的模型的模拟结果,基于器官尺度的模型可以提供关于植物结构的更深入的信息[27]。DSSAT中的测量数据是各类器官的信息,而在GreenLab中,由于可以识别器官序列,还提供了对单个器官生长过程的模拟(图8(a) )以及器官序列的逐个周期的动态变化(图8(b) )。此外,基于构建的GreenLab模型,可以进一步进行计算实验,优化源库参数,在个体尺度上研究结构对产量的影响[16]。
4.2 不同尺度模型结合的应用领域
虚拟农业中的一个研究主题是直观地表达施肥灌溉等措施对作物个体生长的影响。基于本研究的从群体尺度到个体尺度的研究,可以基于模型快速模拟不同管理措施下的作物形态变化,而无需逐个调整器官大小。
5 结 论
本研究通过基于DSSAT中的测量数据和模拟的每日叶、节和果实的重量数据,利用GreenLab模型模拟了四个品种两个处理的玉米植株各个器官的生长发育动态,探索了群体尺度DSSAT与个体尺度GreenLab作物模型连接的可能性。结果表明,基于共同的数据结果,个体尺度的GreenLab模型可和群体尺度的模型输出一致的结果。除了可复现模拟叶面积指数和地上部总重,还可进一步反演出各种器官之间的分配,从而对作物的生长发育进行更详细的描述,并对植物的生长发育过程进行三维可视化展示,从而实现从群体尺度模型到个体尺度模型的转换。





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}